- ÉCOSYSTÈMES
- ÉCOSYSTÈMESL’apport essentiel de l’écologie est sans doute d’avoir montré que les ensembles de populations existant dans un même milieu et présentant entre elles des interactions multiples, telles que relations de cohabitation, de compétition, de prédation, de parasitisme, constituaient avec le milieu où elles vivent des systèmes biologiques, les écosystèmes , qui demandaient une étude globale. Les écosystèmes ont comme tout système une structure qui doit être précisée dans plusieurs domaines. Leur flore comme leur faune possèdent une composition spécifique particulière, affectée d’une variation saisonnière qui caractérise leur structure temporelle. Ces espèces végétales et animales se répartissent en outre dans l’espace en organisant à la fois une structure verticale et une structure en plan. Non moins essentielle est la connaissance de la structure trophique, définie à la fois par la photosynthèse et par les relations de prédation, de parasitisme, de saprophytisme et de saprophagisme entre les espèces: l’écosystème est le siège d’un fonctionnement, lié bien entendu aux structures spatiale et temporelle, et que caractérisent les flux de matière et d’énergie entre ses divers constituants. Il se produit une certaine quantité de matière vivante, la biomasse , qui, par des voies diverses, se transformera en matière organique morte, laquelle sera ensuite progressivement minéralisée en partie ou en totalité.Selon les cas, ce fonctionnement s’inscrit dans un état d’équilibre – une stabilité –, ou au contraire il intervient dans une dynamique évolutive de l’écosystème dont la composition se modifie au cours du temps.Le terme d’écosystème a été introduit par Tansley en 1935 pour qualifier l’ensemble d’une communauté végétale et de son milieu considéré comme une unité. C’était une extension de la notion de biocénose de Möbius (1927) qui ne considérait que les êtres vivants. La notion d’écosystème s’est trouvée précisée ensuite par l’approche tropho-dynamique proposée par Lindeman (1942), lors de l’étude d’un lac aux États-Unis. La biogéocénose de Sukatchev (1947) en est un synonyme, au sens restreint d’une biocénose homogène vivant dans un milieu homogène.Un écosystème peut donc au total être défini comme un système biologique complexe formé par les divers organismes vivant ensemble – une biocénose – dans un milieu donné, et par les éléments de ce milieu qui interviennent dans leur existence – ce que l’on appelle souvent le biotope . Ce fragment de la biosphère constitue une entité relativement autonome par rapport aux écosystèmes voisins et dont il est donc possible – c’est là l’intérêt essentiel du concept – d’en analyser la structure et le fonctionnement.Dans les trois domaines systémiques de la structure, du fonctionnement et de l’équilibre, le milieu physique intervient en liaison étroite avec la biocénose. Lorsque, cependant, on s’intéresse principalement au peuplement constitué par les êtres vivants, on peut en dégager une image assez claire de l’écosystème correspondant, ne serait-ce que par comparaison avec d’autres biocénoses présentes dans des conditions similaires ou dans des conditions différentes. Le lecteur pourra, de ce fait, se reporter à l’article BIOCÉNOSE pour tout ce qui a trait aux structures spatiale, temporelle et trophique de l’écosystème et même, pour une part, à son évolution. Mais le fonctionnement du système, en revanche, ne saurait être dissocié du milieu qui apporte, à l’origine, tous les éléments qui vont constituer la matière vivante, notamment l’énergie, l’eau, le gaz carbonique, les sels minéraux, comme les considérations exposées ci-dessous le démontrent.1. Mesure du fonctionnement d’un écosystèmeBiomasseOn appelle biomasse le poids des êtres vivants appartenant soit à une même espèce, soit à un groupe d’espèces (un groupe ou un niveau trophique par exemple), soit à l’ensemble de la biocénose.L’estimation des biomasses de ses différents constituants donne une vue statique de l’écosystème (fig. 1): leur répartition en niveaux trophiques (cf. ÉCOLOGIE, fig. 9) décrit ainsi quantitativement la structure trophique de l’écosystème. Une biomasse peut être exprimée en poids vif; elle est plus souvent donnée en poids sec (après séchage à 70 0C par exemple, pour éviter les pertes de matière organique qu’entraîneraient des températures plus élevées). Il est préférable de la traduire en contenu d’énergie, c’est-à-dire en kilocalories ou en joules, pour éviter de comptabiliser coquilles et squelettes. Les équivalents énergétiques peuvent être mesurés avec une bombe calorimétrique, mais il est souvent suffisant d’utiliser des tables déjà établies: rappelons qu’à 1 g de glucides correspond en gros 4,1 kcal, à 1 g de protides 4 kcal et à 1 g de lipides 9 kcal.En écologie aquatique, la biomasse peut être rapportée à une unité de volume, centimètre cube ou mètre cube. En écologie terrestre, elle est généralement rapportée à une surface déterminée en mètres carrés, ou en hectares, ou encore en kilomètres carrés selon l’étendue des biocénoses. La figure 1 montre la répartition des biomasses dans le milieu forestier. Remarquons que la biomasse contient, en sus de la matière organique, une certaine quantité de sels minéraux (éléments biogènes ou autres) que l’incinération permet de recueillir sous forme de cendres. Celles-ci sont parfois très importantes dans certaines phytocénoses qui accumulent de la silice et de l’alumine (10 p. 100 de Si2 dans la matière sèche des Equisetum ; 7,1 p. 100 de Al23 dans les feuilles de Symplocos spicata ); la phytocénose peut, dans un tel cas, jouer un rôle géologique important. Duvigneaud (1964) a créé le terme de minéralomasse pour cette fraction de la biomasse qui provient de l’accumulation par les plantes d’éléments minéraux puisés dans le sol par les racines.Après la mort des êtres vivants, la matière organique qui les compose subit une dégradation progressive dont les étapes sont souvent difficiles à distinguer: c’est la litière des feuilles, des branches, des racines, et aussi les cadavres d’animaux et leurs déjections, puis la matière organique non figurée et l’humus du sol. On peut regrouper sous le nom de matière organique morte l’ensemble de ces substances. Son importance par rapport à la matière vivante est très variable selon les écosystèmes et constitue une caractéristique intéressante du système.Des mesures de biomasses (tabl. 1) ont été faites et continuent d’être pratiquées, afin de suivre les effets des mécanismes de dégradation (anthropiques ou physico-chimiques) qui affectent les grands biomes (écosystèmes généraux), puisqu’elles sont, nous l’avons dit, une caractéristique essentielle des écosystèmes. Elles concernent tout spécialement les végétaux de ces écosystèmes, plus faciles à étudier et d’un ordre de grandeur en général très supérieur. Quelques données existent néanmoins aussi sur les zoocénoses. La biomasse des micro-organismes, en revanche, est fort mal connue car difficile à estimer, et par ailleurs de peu de signification par rapport à leur activité métabolique.Flux d’énergie et productivitéLa biomasse ne traduit qu’un aspect statique d’une biocénose, donc d’un écosystème. Plus important est son taux de renouvellement , c’est-à-dire la production biologique – parfois aussi appelée productivité – qui lui correspond (tabl. 1). De manière plus générale, il convient d’étudier les flux d’énergie qui traversent les divers compartiments de l’écosystème et même, plus précisément, tout le réseau trophique. La complexité de ce réseau conduit toutefois à le simplifier considérablement, jusqu’à la représentation schématique extrême que constitue la pyramide des flux d’énergie, où ne figurent que les trois ou quatre niveaux trophiques successifs (fig. 2). On peut généraliser, à partir de cet exemple, en rappelant que les autotrophes (végétaux verts) édifient leur matière organique à partir du gaz carbonique, de l’eau, de nitrates, phosphates et divers autres constituants minéraux, essentiellement par le mécanisme de la photosynthèse que permet l’apport énergétique du rayonnement lumineux. Ils constituent le niveau des producteurs P. Les hétérotrophes, eux, utilisent la matière organique ainsi formée. Ils sont représentés par l’ensemble des animaux, mais aussi par beaucoup de bactéries et par les végétaux non chlorophylliens comme les champignons; ils constituent les niveaux successifs des consommateurs primaires C1 – les herbivores – et secondaires C2 – les carnivores –, niveaux auxquels il convient de rattacher les parasites, les détritivores, les décomposeurs, les fixateurs d’azote.Les difficultés d’une présentation synthétique des flux d’énergie dans un écosystème sont multiples. Il y a d’abord le fait que beaucoup d’espèces sont polyphages et que leur régime alimentaire les place à cheval sur deux ou plusieurs niveaux trophiques: c’est le cas des omnivores et des détritivores. Le cas des micro-organismes est particulièrement complexe, d’abord parce qu’ils se situent aussi à divers niveaux et aussi parce que leur taille microscopique n’en permet pas l’étude selon les mêmes techniques que celles qui concernent les organismes de plus grande taille. Aussi sont-ils souvent absents dans les présentations de flux d’énergie, notamment dans les pyramides des flux d’énergie. On doit ajouter que les micro-organismes ont avec les autres organismes, animaux et végétaux, des relations complexes encore mal connues.Lorsque les divers êtres vivants d’un écosystème auront pu être répartis entre les niveaux trophiques, on pourra faire apparaître comment se distribue le flux d’énergie qui arrive sous forme d’énergie radiante en provenance du Soleil.Le rayonnement du Soleil comporte de l’ultraviolet, mais surtout de l’infrarouge court ( de 0,78 à 3 猪m) et du visible ( de 0,38 à 0,78 猪m). Ces variations sont pour une part réfléchies sur les surfaces de l’écosystème (albédo) et en grande partie utilisées sous forme de chaleur pour combler le déficit thermique et vaporiser les grandes quantités d’eau de transpiration nécessaires au bon fonctionnement des végétaux. Le rayonnement visible fournit la lumière indispensable à la photosynthèse (radiations photosynthétiquement actives, RPA).Du rayonnement lumineux total (LT) qui arrive sur un écosystème, seule une fraction LU frappe la végétation et peut être utilisée (fig. 3), mais de cette fraction une partie seulement sert réellement à l’assimilation chlorophyllienne, l’autre partie, LNA (lumière non assimilée), intervenant dans l’échauffement des plantes et leur transpiration. Le rayonnement utilisé par l’assimilation chlorophyllienne (LA) correspond à ce que l’on appelle souvent production primaire – ou photosynthèse – brute, PB. Elle sert à la synthèse de la matière vivante – production primaire nette ou photosynthèse nette PN –, qui est conjuguée à la respiration des plantes R0 (ou RA, respiration des autotrophes).Les rapports PB/LU et PN/LU sont des rendements physiologiques de photosynthèse – brute et nette – tandis que PB/LT et PN/LT sont les rendements écologiques de la production primaire. C’est sans doute le rendement PN/LT qui rend le mieux compte du rôle des producteurs primaires dans l’écosystème.Au sein de chaque niveau de consommateurs, on trouve ensuite (comme pour toute population), l’énergie ingérée I, l’énergie non assimilée NA, l’énergie assimilée A, elle-même répartie en production P du niveau considéré et énergie R de la respiration à ce niveau. Dans chaque niveau, comme pour une population, le fonctionnement est également caractérisé par les rendements P/I, P/A et A/I, considérés cette fois à l’échelle de l’ensemble du niveau trophique. La représentation graphique utilisée à propos d’une savane africaine dans la figure 4 fait bien apparaître les différences importantes de répartition des flux d’énergie qui existent entre deux niveaux trophiques successifs d’un écosystème.D’un niveau trophique au suivant, il convient de tenir compte de la part non utilisée du niveau supérieur, surtout importante pour la production primaire (NU1), le plus souvent négligeable dans les niveaux de consommateurs (NU2). On peut alors définir les rendements des transferts d’énergie d’un niveau au suivant. Ij /Ij -1 représente ainsi le rendement de l’énergie apportée à un niveau trophique donné j , par exemple I2/I1 pour le passage des consommateurs primaires (herbivores) aux consommateurs secondaires (carnivores), ou I1/LU pour le passage de l’énergie ingérée par les consommateurs primaires à l’énergie lumineuse utilisée par les producteurs chlorophylliens. Aj /Aj -1 est le rendement d’assimilation à un niveau trophique donné et Pj /Pj -1 le rendement de la production à ce niveau: P2/P1 pour les consommateurs secondaires, P1/PN pour les consommateurs primaires. C’est cette succession des valeurs de P dans les niveaux trophiques successifs que traduit graphiquement la classique pyramide des flux d’énergie (fig. 2 à droite).D’autres rendements sont aussi employés par certains auteurs, ce sont les rendements d’utilisation qui font intervenir deux éléments non homologues de niveaux trophiques successifs. Tel est, par exemple, le rapport A2/P1 de l’énergie assimilée par les consommateurs secondaires (carnivores) à l’énergie que représente la production des consommateurs primaires (herbivores).Parmi les points délicats des problèmes de flux d’énergie dans les écosystèmes, il convient de remarquer le fait que les quantités non assimilées NA ne sont pas perdues. Elles seront utilisées par d’autres organismes, coprophages, détritivores et surtout micro-organismes dont le rôle n’a pu jusqu’ici être pleinement pris en compte dans les calculs. De même les quantités d’énergie dites non utilisées NU sont en fait utilisées le plus souvent par des micro-organismes décomposeurs. Ce n’est que sous leur forme respirée (RH = respiration des hétérotrophes) que les énergies sont effectivement perdues par l’écosystème sous forme de chaleur.On appelle parfois productivité nette de l’écosystème PNE la différence entre la productivité d’assimilats par photosynthèse (PB) et tout ce qui est perdu par respiration tant des plantes (RA) que des animaux et micro-organismes (RH):
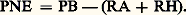 Dans un système en équilibre (climax), la biomasse produite est égale à la quantité consommée, décomposée et minéralisée, d’où: PNE = 0.Le système n’est toutefois pas toujours en équilibre, et une partie de l’énergie fixée est parfois stockée dans l’écosystème. Dans une forêt en croissance, par exemple, un incrément de bois s’ajoute chaque année à la biomasse initiale; dans une toundra, la litière de matière morte ne cesse de s’accroître à cause de l’activité réduite des décomposeurs; dans les tourbières, un humus peu décomposé ajoute chaque année une couche de tourbe à celles des années précédentes. La proportion de matière organique morte par rapport à la matière vivante peut ainsi devenir parfois considérable. C’est à des phénomènes de ce type que nous devons les stocks de lignite, de houille et de pétrole qui se sont accumulés au cours de l’histoire de la Terre.2. Caractéristiques de divers écosystèmes continentauxÉcosystèmes dulçaquicolesLes écosystèmes dulçaquicoles comprennent les milieux limniques, stagnants, et les cours d’eau. Les écosystèmes limniques continentaux – lacs et étangs – ont été parmi les premiers étudiés, car leur relative pauvreté en espèces facilite l’établissement des liens trophiques; par ailleurs, les eaux stagnantes constituent des entités bien délimitées.Dans les écosystèmes limniques, la production primaire, d’origine planctonique, apparaît en général plutôt faible, inférieure à celle des écosystèmes terrestres environnants. C’est plus spécialement le cas, bien entendu, des lacs oligotrophes de montagne.Une seconde caractéristique des écosystèmes aquatiques à dominante planctonique est le fort rendement écologique de croissance des organismes qui constituent le niveau des consommateurs herbivores. La cause en est double: d’une part, un taux d’assimilation élevé en rapport avec la faible proportion de cellulose dans le phytoplancton – et inversement sa forte teneur en protéines; d’autre part, le fait que les consommateurs sont tous des hétérothermes, qu’ils soient crustacés, rotifères ou même poissons, et jamais des vertébrés homéothermes. Le phytoplancton étant presque complètement consommable par les herbivores planctoniques, cela diminue l’importance des chaînes partant des détritivores – dont le rendement est toujours faible – ainsi que la quantité de matière végétale «non utilisée», c’est-à-dire en fait décomposée par les micro-organismes: la proportion de matière végétale ainsi consommée à l’état vivant atteint 90 p. 100 dans le cas du phytoplancton d’un lac oligotrophe contre bien moins de 10 p. 100 dans une forêt.La faible part de la végétation «non utilisée» jointe au fort rendement écologique de croissance des herbivores conduisent dans les milieux aquatiques à des rendements de transfert élevés du niveau des producteurs à celui de leurs consommateurs: ces rendements atteignent ainsi des valeurs de l’ordre de 0,05 à 0,15 et parfois même davantage (fig. 2). En revanche, entre le niveau des herbivores et celui des carnivores, les transferts d’énergie ne sont pas très différents de ce qu’ils sont dans les écosystèmes terrestres: les exemples étudiés les révèlent compris entre 0,05 et 0,22. Mais, l’originalité des milieux limniques réapparaît si on considère le taux de renouvellement de la biomasse, qui fait intervenir le facteur temps dans les transferts d’énergie. Dans le milieu aquatique dominent, en effet, des espèces de petite taille – copépodes, rotifères, cladocères par exemple – et aussi des organismes unicellulaires, notamment le phytoplancton, dont la vie moyenne est particulièrement courte. Il en découle que le renouvellement de la biomasse est nettement plus rapide dans les écosystèmes aquatiques que dans les écosystèmes terrestres. Le fait est particulièrement net pour le niveau des producteurs. Là, comme nous l’avons signalé, un rôle souvent dominant revient aux organismes microscopiques du phytoplancton, dont la durée de vie ne dépasse pas quelques heures en période chaude et, même en période froide, n’excède que rarement une ou quelques semaines. Les taux de renouvellement annuels P/B atteignent alors et dépassent même des valeurs de plusieurs dizaines.On est loin évidemment des valeurs observées pour les plantes des milieux terrestres, qui sont de l’ordre de 1 pour les herbes, et descendent souvent à bien moins de 0,1 pour les arbres, correspondant ainsi à des durées de vie moyenne de l’ordre de 1 an à plus de 30 ans, respectivement.Le taux de renouvellement élevé de la biomasse des producteurs se traduit par une pyramide des biomasses à base relativement étroite par rapport à celle des écosystèmes herbacés et plus encore forestiers. Il ne faut toutefois jamais perdre de vue que les considérations qui précèdent ne concernent que les éléments visibles et facilement dénombrables des biocénoses. Les micro-organismes, qu’il s’agisse des bactéries ou des protozoaires, ne sont le plus souvent pas pris en compte, ce qui, évidemment, fausse grandement la conception de niveau trophique. Dans la réalité, ces micro-organismes jouent pourtant un rôle essentiel dès le niveau des consommateurs primaires: les protozoaires, en se nourrissant de phytoplancton, et surtout les bactéries, en utilisant la matière organique et la cellulose des végétaux. Le rôle de ces dernières sera donc d’autant plus important que les végétaux comporteront une part plus grande non ou difficilement assimilable par les herbivores, ce qui est le cas essentiellement des végétaux terrestres et, en milieu aquatique, des macrophytes.De fait, toutes les remarques faites jusqu’ici se rapportent essentiellement aux milieux limniques où la production primaire est due au phytoplancton. Tel n’est plus le cas lorsque la profondeur est très faible et que des macrophytes, tels que nénuphars, massettes, roseaux, prennent une part importante dans la production primaire. La situation se rapproche alors de celle des milieux terrestres herbacés, car ces végétaux ne sont que très partiellement consommés par les herbivores. La production primaire de tels écosystèmes est plus élevée que celle des écosystèmes à dominante planctonique, mais les chaînes de détritivores deviennent prépondérantes, avec un mauvais rendement écologique de croissance et un taux de renouvellement faible si l’on ne considère pas les bactéries.On peut remarquer que les écosystèmes d’eaux courantes tant soit peu rapides, qui ne permettent pas le développement du plancton et qui, par ailleurs, peuvent recevoir, en provenance des écosystèmes terrestres environnants, des apports importants de débris végétaux riches en cellulose et en lignine, se comportent un peu, à certains points de vue, comme des chaînes de détritivores.Savanes tropicalesLes milieux de ce type, caractérisés par la présence d’un tapis herbacé dense parsemé de formes ligneuses, arbres ou arbustes, occupent dans le monde de vastes superficies. Elles sont liées au monde tropical où alternent deux saisons contrastées, l’une pluvieuse, l’autre sèche. Leur structure et surtout leur fonctionnement dépendent étroitement de la quantité de pluie et de sa répartition saisonnière.Dans la savane préforestière de Lamto, en Côte-d’Ivoire, milieu herbacé parsemé d’arbres, d’arbustes et de palmiers, la biomasse peut atteindre en matière sèche (MS) 66 t/ha, dont 42 épigées, tandis que la production primaire est de 28,5 t/ha/an (114 憐 106 kcal/ha/an) dont 15 épigées. Bien entendu, la part relative des herbes et celle des arbres et des palmiers est très différente selon qu’il s’agit des biomasses ou des productions: avec une biomasse bien moins importante que celle des plantes ligneuses, les herbes assurent une production bien supérieure.Le schéma de la figure 4 traduit les flux d’énergie dans les organismes non microscopiques des niveaux trophiques successifs de cette savane.De la production primaire P = 114 憐 106 kcal/ha/an, seule une fraction (I = 62 憐 106 kcal/ha/an) est consommée par les animaux. Une autre fraction est brûlée chaque année par le feu de brousse (8 t/ha/an soit 36 憐 106 kcal/ha/an) et une troisième partie (NU = 16 憐 106 kcal/ha/an) semble non utilisée. Il est apparu également que l’énergie ingérée par les consommateurs primaires n’était que pour une très faible part réellement assimilée: plus de 90 p. 100 est rejetée sous forme d’excréments (NA = 54 憐 106 kcal/ha/an). Ce faible rendement d’assimilation est lié à la dominance des vers de terre. Plus avant dans le fonctionnement, au niveau des consommateurs secondaires C2 et C3 – les carnivores –, les rendements d’assimilation A/I et de production biologique P/I deviennent au contraire excellents (fig. 5).On a montré par ailleurs que les micro-organismes consommaient dans le sol 54 憐 106 kcal/ha/an soit l’équivalent de 13 t/ha/an, et 12 憐 106 kcal/ha/an, soit environ 3 t/ha/an au-dessus du sol (début de décomposition des feuilles). Cette activité (NU) correspond précisément à l’énergie en apparence non utilisée et à l’énergie non assimilée par les herbivores (NA). Dans les savanes plus sèches, au Sahel par exemple, la production primaire diminue progressivement. En revanche, la part relative consommée par les organismes non microscopiques est plus importante, par suite sans doute d’un ralentissement des phénomènes de décomposition bactérienne lié à la sécheresse. Les fourmis et les termites jouent un rôle de plus en plus essentiel, tandis que les vers de terre perdent de leur importance.ForêtsForêts tempérées caducifoliéesCe sont les écosystèmes les plus étudiés, tant en Amérique du Nord (érablière, tuliperaie) qu’en Europe (chênaie, hêtraie). Dans les forêts tempérées caducifoliées, la phytomasse totale atteint en fin de développement (climax) des valeurs de 250 à 450 t MS (matière sèche)/ha (fig. 1 et tabl. 1). Les racines en représentent de 15 à 20 p. 100 et le poids des feuilles reste de l’ordre de 3 à 4 t MS/ha.La productivité brute PB, qui se mesure par les méthodes sophistiquées de l’écophysiologie, est approximativement le double de la productivité primaire nette PN. Cette productivité a été mesurée en de nombreux endroits; lors de l’optimum de croissance de la phytocénose, elle est de l’ordre de 11 à 16 t MS/ha/an, ce qui correspond à 45-70.106 kcal/ha/an, et diminue ensuite progressivement.Au cours d’une année, la période de végétation (phénophase feuillée) n’est que de 4 à 6 mois et la productivité primaire se partage ainsi en :– une fraction qui meurt et tombe, surtout à l’automne, sous forme de chute de litière utilisée par des détritivores;– un apport aux consommateurs herbivores ou phytophages;– une exportation de matériaux divers, surtout importante lorsqu’il y a exploitation par l’homme (éclaircies ou coupes à blanc);– une contribution à l’augmentation de la phytomasse ligneuse: c’est l’incrément de la forêt souvent exprimé en mètres cubes de bois fort (jusqu’à 7 cm de diamètre) par les forestiers occidentaux. Cet incrément est de 6-13 t MS/ha, soit 3 à 8 m3 à l’hectare de bois fort pour le hêtre et environ 5 m3 pour le chêne, dont la densité est plus forte; pour des bois légers à croissance rapide comme celui de certains résineux, l’incrément peut être bien plus important: il atteint jusqu’à 20 m3 pour l’épicéa.La fraction de la production nette PN entrant dans le réseau de consommation par l’intermédiaire des herbivores et phytophages est généralement très faible; les vertébrés sont peu importants: 5-7 kg MS/ha pour les mammifères, 1-2 kg MS/ha pour les oiseaux. Les invertébrés peuvent être bien plus abondants, surtout lors d’années d’invasion par des chenilles défoliatrices qui affectent profondément la fixation d’énergie: leur biomasse peut alors passer de 10-20 kg MS/ha (valeur normale) à 75-150 kg MS/ha, ce qui correspond à une destruction de plus de la moitié de la production foliaire. L’entomofaune circulante, qui atteint une biomasse de 15-20 kg MS/ha, est composée d’insectes (surtout diptères) commençant leur vie à l’état de larve dans la litière et dans l’humus (rôles sapro-, rhizo- ou zoophages). Tous ces animaux n’utilisent qu’une partie de la phytomasse consommée à édifier leur biomasse (productivité secondaire); le reste est perdu sous forme de fèces et d’énergie respiratoire.La chaîne des décomposeurs l’emporte totalement sur celle des consommateurs. En effet 50 à 75 p. 100 de la productivité primaire nette donnent chaque année une litière qui entre dans le réseau de décomposition. De cette litière une fraction, lente à se décomposer, subsistera pendant un nombre plus ou moins grand d’années, s’ajoutant à la litière préexistante; une autre fraction, livrée aux détritivores et aux transformateurs, deviendra de l’humus; le restant sera reminéralisé (recyclage) par des bactéries minéralisatrices.La biomasse des bactéries n’est que de quelques kilogrammes de matière sèche par hectare, mais leur productivité est particulièrement forte, car elles ne cessent de se multiplier en même temps qu’elles manifestent les activités les plus diverses – décomposition, nitrification, dénitrification, fixation de l’azote, reminéralisation. Les animaux décomposeurs de la pédofaune sont avant tout des bactério et mycophages pâturant les peuplements de micro-organismes développés dans et sur la litière.Forêts de résineuxLes grandes forêts boréales toujours vertes de l’hémisphère Nord, adaptées à des climats souvent difficiles, n’ont en zone de taïga qu’une productivité médiocre. Rodin et Basilevic donnent, pour les forêts de l’U.R.S.S., les valeurs moyennes suivantes :– biomasse en tonnes par hectare = 100 en taïga du Nord, 260 en taïga centrale et 330 en taïga du Sud;– productivité nette par hectare et par an, respectivement 4,5 t, 7 t et 8,5 t.Cependant, la productivité varie fortement avec l’âge et, dans la taïga du Sud, elle peut passer par un pic de 20 t/ha/an pour les forêts en pleine croissance sur bon sol, la moyenne étant de 14 t/ha/an.L’épicéa a été et est encore aujourd’hui la principale essence de reboisement des zones peu fertiles de l’Europe occidentale. Ici, protégées par l’homme, les plantations monospécifiques d’épicéas (écosystèmes culturaux) réagissent bien mieux à un climat plus favorable que celui de leur pays d’origine, et leur production nette atteint 15-20 t/ha/an, même sur des sols médiocres.Les pins (Pinus) , grâce à leur grande frugalité, colonisent des sols très pauvres. Ainsi, la principale «usine à bois» de l’Europe occidentale est la forêt de pins maritimes (Pinus pinaster) rétablie dans le triangle des landes de Gascogne. Entre 24 et 28 ans, l’accroissement des troncs atteint de 5 à 20 m3/ha et la production globale est de l’ordre de 14 t/ha/an.Forêts tropicalesLes forêts tropicales, particulièrement les forêts tropicales humides (forêts sempervirentes ou «rain forest»), sont caractérisées par la luxuriance de leur végétation dans laquelle se superposent une série de strates; les arbres les plus hauts peuvent dépasser 50 m.Une solide étude d’écosystème forestier tropical humide a été réalisée en Malaisie à Pasow. La biomasse épigée dépasse 430 t (matière sèche) par hectare, dont 346 t pour les troncs et moins de 8 t pour les feuilles. La production primaire nette peut être estimée à 26,7 t/ha/an, dont 13,7 concernent les parties ligneuses et 7,4 les feuilles. La respiration de ces végétaux est intense, par suite de la température constamment élevée, même la nuit: elle représente l’équivalent de 50,5 t/ha/an, ce qui permet d’estimer la production brute de l’écosystème à 77,2 t/ha/an. L’écart considérable entre production brute et production nette est caractéristique des milieux tropicaux, et tout spécialement des forêts sempervirentes.Sur cette production, la quantité consommée vivante est excessivement faible: de l’ordre de 0,3 t/ha/an seulement, ce qui signifie que l’essentiel de la matière vivante formée est utilisé après sa mort par des détritivores et plus encore par des micro-organismes décomposeurs. Quant à la grande faune des forêts, si elle peut être très diversifiée, elle n’est représentée que par de très faibles densités d’individus de chaque espèce. De plus, elle est en grande partie dans les strates supérieures (singes, oiseaux), là précisément où se trouvent les feuilles vivantes qui sont le point de départ des chaînes trophiques.ToundrasTout à l’opposé des forêts tropicales, les toundras sont des formations végétales très basses localisées dans les régions les plus froides du globe. Les formes végétales dominantes y sont des lichens ou des mousses.La production annuelle est très faible, inférieure à 3 t/ha/an, et le temps de renouvellement de la biomasse relativement très long pour des plantes herbacées de petite taille. La vitesse de décomposition est également très lente.La vie animale, surtout endogée, et pour une part aquatique, dans un sol gelé une grande partie de l’année, mais imbibé d’eau, est représentée surtout par des détritivores dont l’action est complétée par celle des micro-organismes décomposeurs qui ont, par suite du froid, une action très ralentie. De fait, le cycle saisonnier est très accentué et marqué par une longue phase d’inactivité hivernale. Quelques vertébrés supérieurs peuvent survivre, à la faveur de l’homéothermie, mais leur importance quantitative est très faible, qu’ils soient herbivores ou carnivores.Prairies des zones tempéréesContrairement aux écosystèmes forestiers, la prairie est destinée à développer la productivité secondaire des grands herbivores. Relativement appréciable, celle-ci sera «exportée» sous forme de viande, de lait, de cuir, de laine. Ce sont pourtant encore ici les décomposeurs et reminéralisateurs (lombrics, bactéries, champignons) qui profitent le plus d’une productivité primaire où la part des racines est très importante.Cette prairie peut devenir permanente, ou faire partie d’un système de rotation dans lequel elle est périodiquement retournée pour faire place à des cultures. La récolte de l’herbe – surtout graminées et légumineuses – se fait de deux façons: la fauche (prés à foin) ou la dent des animaux (pâtures), ou parfois des deux combinées.La productivité primaire dépend du nombre de fauches ou du nombre de passages du bétail; en fonction de ce nombre, elle augmente d’abord, pour diminuer ensuite (surpâturage destructeur). Elle est en général de 5 à 15 t MS/ha/an pour les parties aériennes, mais irrigation et engrais peuvent, dans quelques cas, l’amener à 25 t. La productivité des racines est difficile à déterminer, celles-ci se renouvelant tous les 1, 2, 3 ans ou plus, selon les espèces; de plus, les racines sont des organes régulateurs, participant à la repousse de l’herbe après chaque coupe, grâce aux matières stockées pendant les intervalles. La phytomasse des racines est de l’ordre de 10 t MS/ha et l’on peut en estimer la productivité à 5 t/ha ou même davantage.Les animaux pâturants ne sont sur le terrain que pendant la période de végétation active (180-225 j/an); leur productivité dépend de la quantité et de la qualité de l’herbe (digestibilité, protéines digestibles, oligoéléments); elle dépend aussi du nombre de passages (division de la prairie en parcelles). On a établi que, dans de bonnes conditions, une vache laitière de 500 kg mangeant 70 kg d’herbe (14 kg MS par jour) produit 15 l de lait par jour; un bœuf à l’engrais de 350 kg accroît son poids vif de 1 kg/j en mangeant 45 kg d’herbe (9 kg MS) par jour.Le rendement et la saison de pâturage peuvent être augmentés par l’utilisation de foin et d’aliments plus concentrés. Ces aliments fournis à l’étable sont souvent produits à la ferme par des écosystèmes annexés à la prairie permanente: prés de fauches, luzernières, cultures de céréales ou plantes à tubercules, etc. Cela nous introduit dans un type d’écosystème plus complexe, l’agroécosystème.AgroécosystèmesAu sens large, l’agroécosystème est le plus ancien écosystème étudié et mis en place par les hommes du milieu rural. Une fois réalisée la domestication des plantes et des animaux, il devenait vital pour les paysans d’obtenir les meilleurs rendements de leur terre, en assurant les assolements nécessaires, la conservation de l’humus et le recyclage des nutriments minéraux par l’équilibre agro-pastoral .La forêt joue aussi un rôle dans la valorisation des terres agricoles (matériaux divers, bois de feu, haies pare-vent, pâturage sous forêt, etc.), de sorte que l’on peut parler, avec Kuhnholz-Lordat, d’équilibre sylvo-agro-pastoral . Il s’agit de maintenir, suivant les régions, des proportions équilibrées écologiquement de forêts (silva ), de pâturages (saltus ) et de cultures (ager ); la localisation topographique peut être aussi très importante: les forêts sur les crêtes régularisent le cycle de l’eau et empêchent l’inondation des vallons, où les apports réguliers d’alluvions fertiles forment des sols favorables aux cultures; les pentes, où le travail de la terre est plus difficile, peuvent servir de pâturages, à condition d’en éviter la surcharge.On voit apparaître ici les notions de paysage écologiquement fonctionnel, d’écopaysage, voire d’écorégion.Ayant envisagé déjà les cas de la forêt et de la prairie pâturée, nous considérerons ici l’agroécosystème dans son sens étroit de champ cultivé. Une partie très importante de la productivité primaire est exportée, non seulement par la récolte des grains, tubercules ou autres organes recherchés, mais souvent aussi sous forme de paille, feuilles ou déchets divers servant à l’alimentation animale, à la litière d’écurie, au compostage, etc. C’est dire que la restitution au sol de matière organique et de nutriments minéraux est faible, ce qui nécessite une importation compensant les matières premières perdues si l’on veut éviter une dégradation et un appauvrissement progressif du sol; aujourd’hui, le traditionnel fumier fait une place de plus en plus grande aux engrais chimiques.Pour fixer les idées, la productivité primaire nette d’un bon champ de blé, sur sol sablo-limoneux fertile, est de 16,7 t MS/ha, dont 6,5 t de grains, 2,2 t de glumes et glumelles, 5 t de tiges, 2 t de feuilles, 1 t de racines. Les 6,5 t de grains et 7,5 t de «pailles» (glumes, tiges et feuilles séchées) sont exportées; 2,7 t de bas de tiges, racines et feuilles sont restituées, sous forme d’éteule abandonnée sur le terrain après la coupe.Dans les pays développés, la productivité des cultures est très fortement augmentée par l’importation de grandes quantités d’énergie fossile (pétrole, gaz naturel) nécessaire (machines agricoles, transports, irrigation, engrais, pesticides, aménagement des installations, etc.).L’exemple de l’écosystème «champ de maïs» aux Etats-Unis, étudié par Pimentel et ses collaborateurs (1973) montre que, en moyenne, le champ de maïs reçoit, pendant la période de végétation, 5,1.109 kilocalories d’énergie solaire totale, par hectare; 1,26 p. 100 de cette énergie est convertie en production (PN1), de maïs total, soit 64 360.103 kcal, correspondant à 16 t MS/ha/an; environ le tiers de cette production correspond aux grains: 20 432.103 kcal (5,1 t MS grains/ha/an), représentant en tout environ 0,4 p. 100 de l’énergie solaire reçue (fig. 6). Une telle productivité ne peut être obtenue qu’avec l’intervention, à de très nombreux niveaux, de quantités importantes de combustibles fossiles:– énergie de fabrication des engrais (h), surtout azotés: en tout 2 641.103 kcal/ha;– énergie de fabrication des pesticides (j);– énergie d’irrigation (i): en moyenne, 85.103 kcal d’énergie sont nécessaires pour amener l’eau et la distribuer sur 1 hectare de culture de maïs;– énergie de séchage car le grain mûr de maïs hybride qui contient 13 p. 100 d’eau risque de moisir et il faut utiliser 300.103 kcal d’énergie pour le sécher;– énergie électrique pour les besoins les plus variés de la ferme (b, c): 775.103 kcal pour 1 hectare de champ cultivé;– apport de graines sélectionnées (maïs hybride) pour plantation, soit une importation d’énergie dans le système de 157.503 kcal;– énergie humaine (d): aux États-Unis où un ouvrier agricole travaille 40 heures par semaine en consommant 21 770 kcal/semaine et où il passe, pour toute la saison, 22 h sur 1 hectare de champ de maïs, cela représente 12,3.103 kcal d’énergie alimentaire.Ces divers apports d’énergie représentent plus du tiers de l’énergie solaire fixée sous forme de grains (1 kcal d’énergie fossile pour produire 2,8 kcal de grains). Dans les cas, très fréquents, où les grains de maïs servent à l’alimentation des animaux domestiques, il faut dès lors compter 3 kcal d’énergie fossile pour produire 1 kcal de viande par l’intermédiaire de 9 kcal de grains.Si l’on considère, par ailleurs, que l’alimentation humaine est à la fois d’origine végétale et d’origine animale, c’est-à-dire que l’homme se situe à la fois au niveau des herbivores et à celui des carnivores et qu’entre ces deux niveaux la perte d’énergie est de 100 à 1, on voit d’après l’exemple précédent quelle économie énergétique peut représenter pour l’humanité de demain un régime de plus en plus végétarien. De fait, les populations qui doivent vivre à des densités considérables, comme c’est le cas des habitants des deltas indochinois par exemple, ont depuis toujours une alimentation essentiellement végétale.3. Contrôle du fonctionnementÀ l’inverse d’un organisme, un écosystème ne possède pas d’organes de contrôle dont le rôle serait analogue à celui des systèmes neuro-endocrinien et immunitaire des vertébrés, dont la fonction est d’assurer à la fois l’intégration du fonctionnement des organes effecteurs, la constance du milieu intérieur et l’adaptation optimale aux conditions changeantes de l’environnement. L’écosystème n’est donc pas une sorte de «superorganisme» installé dans un ensemble de conditions physiques qui lui conviennent. La cohésion de sa composante vivante est entièrement assurée par la complémentarité d’intérêts (en termes de valeur adaptative, ou fitness ) des espèces qui la composent. Celles-ci sont associées sous l’effet de deux types de contraintes, les unes externes, les autres internes.Les contraintes externes constituent ce que l’on appelle couramment les facteurs limitants du système, dont ils déterminent à la fois les limites spatiales et la structure temporelle; ils influencent aussi le taux de renouvellement de ses composantes vivantes. L’identification des principaux facteurs limitants des écosystèmes est ancienne, bien antérieure à la naissance de l’écologie en tant que discipline autonome. Dans les écosystèmes terrestres, les rôles capitaux de la température et des précipitations, de l’altitude et de la latitude, ont été reconnus dès le début du XIXe siècle; les lignes isothermes, les isohyètes et les courbes de niveau interviennent à ce titre dans tous les travaux des biogéographes. La lumière ou la teneur en oxygène et en sels minéraux jouent un rôle essentiel dans les milieux aquatiques. La restriction spatiale et l’isolement géographique ont depuis longtemps été impliqués en tant que variables influençant la richesse spécifique et par conséquent la structure trophique des biocénoses.L’analyse des modalités d’action de ces facteurs limitants exogènes est cependant bien loin d’être achevée. On a, par exemple, trop utilisé jusqu’ici les moyennes annuelles de la température ou des précipitations, et trop souvent sous-estimé les valeurs extrêmes. Quelle que soit la latitude, en effet, les variations saisonnières des paramètres climatiques majeurs ont de profondes répercussions sur la structure et la dynamique des écosystèmes. L’importance des variations journalières des mêmes variables n’a pas reçu non plus toute l’attention qu’elle mérite, en particulier sur les hautes montagnes tropicales. Dans les phénomènes d’insularité, l’importance de la distance des sources possibles de migration et des phénomènes d’extinction spontanée n’a été reconnue que très récemment.À côté de ces contraintes exogènes, la part des contraintes endogènes est difficile à apprécier dans un écosystème naturel «mature» dont l’homéostasie est généralement excellente. En revanche, dès que l’on considère des successions dans le temps (séries biocénotiques progressives ou régressives) ou certaines situations expérimentales (agrosystèmes), il devient possible d’entrevoir l’importance de certains mécanismes internes de régulation. Les uns ont pour fonction de limiter la taille des populations spécifiques, de façon à éviter qu’elles ne dépassent la capacité de charge du milieu: tel est le cas des facteurs socioécologiques de régulation de la natalité et de la mortalité dépendant de la densité, dont l’existence semble certaine chez un certain nombre de vertébrés. Quand ces facteurs ne jouent plus, généralement du fait d’une intervention humaine, une pullulation de consommateurs peut aboutir à la destruction de leur propre habitat: la déforestation quasi totale de certains parcs nationaux africains consécutive à une surpopulation par les éléphants est un exemple connu de tous.D’autres mécanismes internes de régulation sont interspécifiques et ont pour effet d’empêcher ou de limiter la compétition à un niveau trophique donné. Telle paraît être la fonction de ces substances chimiques sécrétées par beaucoup de végétaux et qui peuvent empêcher la croissance d’espèces concurrentes (phénomènes d’antibiose et d’allélopathie). Certains agents pathogènes animaux (des virus aux hématozoaires) qui ne causent que des accidents mineurs chez leurs hôtes habituels, mais peuvent entraîner une mortalité très élevée chez des formes voisines non sympatriques, ont peut-être un rôle écologique voisin (myxomatose, herpès simien, certains trypanosomes, etc.). Des phéromones animales, en particulier celles exerçant un pouvoir répulsif, sont peut-être à ranger aussi dans cette catégorie.
Dans un système en équilibre (climax), la biomasse produite est égale à la quantité consommée, décomposée et minéralisée, d’où: PNE = 0.Le système n’est toutefois pas toujours en équilibre, et une partie de l’énergie fixée est parfois stockée dans l’écosystème. Dans une forêt en croissance, par exemple, un incrément de bois s’ajoute chaque année à la biomasse initiale; dans une toundra, la litière de matière morte ne cesse de s’accroître à cause de l’activité réduite des décomposeurs; dans les tourbières, un humus peu décomposé ajoute chaque année une couche de tourbe à celles des années précédentes. La proportion de matière organique morte par rapport à la matière vivante peut ainsi devenir parfois considérable. C’est à des phénomènes de ce type que nous devons les stocks de lignite, de houille et de pétrole qui se sont accumulés au cours de l’histoire de la Terre.2. Caractéristiques de divers écosystèmes continentauxÉcosystèmes dulçaquicolesLes écosystèmes dulçaquicoles comprennent les milieux limniques, stagnants, et les cours d’eau. Les écosystèmes limniques continentaux – lacs et étangs – ont été parmi les premiers étudiés, car leur relative pauvreté en espèces facilite l’établissement des liens trophiques; par ailleurs, les eaux stagnantes constituent des entités bien délimitées.Dans les écosystèmes limniques, la production primaire, d’origine planctonique, apparaît en général plutôt faible, inférieure à celle des écosystèmes terrestres environnants. C’est plus spécialement le cas, bien entendu, des lacs oligotrophes de montagne.Une seconde caractéristique des écosystèmes aquatiques à dominante planctonique est le fort rendement écologique de croissance des organismes qui constituent le niveau des consommateurs herbivores. La cause en est double: d’une part, un taux d’assimilation élevé en rapport avec la faible proportion de cellulose dans le phytoplancton – et inversement sa forte teneur en protéines; d’autre part, le fait que les consommateurs sont tous des hétérothermes, qu’ils soient crustacés, rotifères ou même poissons, et jamais des vertébrés homéothermes. Le phytoplancton étant presque complètement consommable par les herbivores planctoniques, cela diminue l’importance des chaînes partant des détritivores – dont le rendement est toujours faible – ainsi que la quantité de matière végétale «non utilisée», c’est-à-dire en fait décomposée par les micro-organismes: la proportion de matière végétale ainsi consommée à l’état vivant atteint 90 p. 100 dans le cas du phytoplancton d’un lac oligotrophe contre bien moins de 10 p. 100 dans une forêt.La faible part de la végétation «non utilisée» jointe au fort rendement écologique de croissance des herbivores conduisent dans les milieux aquatiques à des rendements de transfert élevés du niveau des producteurs à celui de leurs consommateurs: ces rendements atteignent ainsi des valeurs de l’ordre de 0,05 à 0,15 et parfois même davantage (fig. 2). En revanche, entre le niveau des herbivores et celui des carnivores, les transferts d’énergie ne sont pas très différents de ce qu’ils sont dans les écosystèmes terrestres: les exemples étudiés les révèlent compris entre 0,05 et 0,22. Mais, l’originalité des milieux limniques réapparaît si on considère le taux de renouvellement de la biomasse, qui fait intervenir le facteur temps dans les transferts d’énergie. Dans le milieu aquatique dominent, en effet, des espèces de petite taille – copépodes, rotifères, cladocères par exemple – et aussi des organismes unicellulaires, notamment le phytoplancton, dont la vie moyenne est particulièrement courte. Il en découle que le renouvellement de la biomasse est nettement plus rapide dans les écosystèmes aquatiques que dans les écosystèmes terrestres. Le fait est particulièrement net pour le niveau des producteurs. Là, comme nous l’avons signalé, un rôle souvent dominant revient aux organismes microscopiques du phytoplancton, dont la durée de vie ne dépasse pas quelques heures en période chaude et, même en période froide, n’excède que rarement une ou quelques semaines. Les taux de renouvellement annuels P/B atteignent alors et dépassent même des valeurs de plusieurs dizaines.On est loin évidemment des valeurs observées pour les plantes des milieux terrestres, qui sont de l’ordre de 1 pour les herbes, et descendent souvent à bien moins de 0,1 pour les arbres, correspondant ainsi à des durées de vie moyenne de l’ordre de 1 an à plus de 30 ans, respectivement.Le taux de renouvellement élevé de la biomasse des producteurs se traduit par une pyramide des biomasses à base relativement étroite par rapport à celle des écosystèmes herbacés et plus encore forestiers. Il ne faut toutefois jamais perdre de vue que les considérations qui précèdent ne concernent que les éléments visibles et facilement dénombrables des biocénoses. Les micro-organismes, qu’il s’agisse des bactéries ou des protozoaires, ne sont le plus souvent pas pris en compte, ce qui, évidemment, fausse grandement la conception de niveau trophique. Dans la réalité, ces micro-organismes jouent pourtant un rôle essentiel dès le niveau des consommateurs primaires: les protozoaires, en se nourrissant de phytoplancton, et surtout les bactéries, en utilisant la matière organique et la cellulose des végétaux. Le rôle de ces dernières sera donc d’autant plus important que les végétaux comporteront une part plus grande non ou difficilement assimilable par les herbivores, ce qui est le cas essentiellement des végétaux terrestres et, en milieu aquatique, des macrophytes.De fait, toutes les remarques faites jusqu’ici se rapportent essentiellement aux milieux limniques où la production primaire est due au phytoplancton. Tel n’est plus le cas lorsque la profondeur est très faible et que des macrophytes, tels que nénuphars, massettes, roseaux, prennent une part importante dans la production primaire. La situation se rapproche alors de celle des milieux terrestres herbacés, car ces végétaux ne sont que très partiellement consommés par les herbivores. La production primaire de tels écosystèmes est plus élevée que celle des écosystèmes à dominante planctonique, mais les chaînes de détritivores deviennent prépondérantes, avec un mauvais rendement écologique de croissance et un taux de renouvellement faible si l’on ne considère pas les bactéries.On peut remarquer que les écosystèmes d’eaux courantes tant soit peu rapides, qui ne permettent pas le développement du plancton et qui, par ailleurs, peuvent recevoir, en provenance des écosystèmes terrestres environnants, des apports importants de débris végétaux riches en cellulose et en lignine, se comportent un peu, à certains points de vue, comme des chaînes de détritivores.Savanes tropicalesLes milieux de ce type, caractérisés par la présence d’un tapis herbacé dense parsemé de formes ligneuses, arbres ou arbustes, occupent dans le monde de vastes superficies. Elles sont liées au monde tropical où alternent deux saisons contrastées, l’une pluvieuse, l’autre sèche. Leur structure et surtout leur fonctionnement dépendent étroitement de la quantité de pluie et de sa répartition saisonnière.Dans la savane préforestière de Lamto, en Côte-d’Ivoire, milieu herbacé parsemé d’arbres, d’arbustes et de palmiers, la biomasse peut atteindre en matière sèche (MS) 66 t/ha, dont 42 épigées, tandis que la production primaire est de 28,5 t/ha/an (114 憐 106 kcal/ha/an) dont 15 épigées. Bien entendu, la part relative des herbes et celle des arbres et des palmiers est très différente selon qu’il s’agit des biomasses ou des productions: avec une biomasse bien moins importante que celle des plantes ligneuses, les herbes assurent une production bien supérieure.Le schéma de la figure 4 traduit les flux d’énergie dans les organismes non microscopiques des niveaux trophiques successifs de cette savane.De la production primaire P = 114 憐 106 kcal/ha/an, seule une fraction (I = 62 憐 106 kcal/ha/an) est consommée par les animaux. Une autre fraction est brûlée chaque année par le feu de brousse (8 t/ha/an soit 36 憐 106 kcal/ha/an) et une troisième partie (NU = 16 憐 106 kcal/ha/an) semble non utilisée. Il est apparu également que l’énergie ingérée par les consommateurs primaires n’était que pour une très faible part réellement assimilée: plus de 90 p. 100 est rejetée sous forme d’excréments (NA = 54 憐 106 kcal/ha/an). Ce faible rendement d’assimilation est lié à la dominance des vers de terre. Plus avant dans le fonctionnement, au niveau des consommateurs secondaires C2 et C3 – les carnivores –, les rendements d’assimilation A/I et de production biologique P/I deviennent au contraire excellents (fig. 5).On a montré par ailleurs que les micro-organismes consommaient dans le sol 54 憐 106 kcal/ha/an soit l’équivalent de 13 t/ha/an, et 12 憐 106 kcal/ha/an, soit environ 3 t/ha/an au-dessus du sol (début de décomposition des feuilles). Cette activité (NU) correspond précisément à l’énergie en apparence non utilisée et à l’énergie non assimilée par les herbivores (NA). Dans les savanes plus sèches, au Sahel par exemple, la production primaire diminue progressivement. En revanche, la part relative consommée par les organismes non microscopiques est plus importante, par suite sans doute d’un ralentissement des phénomènes de décomposition bactérienne lié à la sécheresse. Les fourmis et les termites jouent un rôle de plus en plus essentiel, tandis que les vers de terre perdent de leur importance.ForêtsForêts tempérées caducifoliéesCe sont les écosystèmes les plus étudiés, tant en Amérique du Nord (érablière, tuliperaie) qu’en Europe (chênaie, hêtraie). Dans les forêts tempérées caducifoliées, la phytomasse totale atteint en fin de développement (climax) des valeurs de 250 à 450 t MS (matière sèche)/ha (fig. 1 et tabl. 1). Les racines en représentent de 15 à 20 p. 100 et le poids des feuilles reste de l’ordre de 3 à 4 t MS/ha.La productivité brute PB, qui se mesure par les méthodes sophistiquées de l’écophysiologie, est approximativement le double de la productivité primaire nette PN. Cette productivité a été mesurée en de nombreux endroits; lors de l’optimum de croissance de la phytocénose, elle est de l’ordre de 11 à 16 t MS/ha/an, ce qui correspond à 45-70.106 kcal/ha/an, et diminue ensuite progressivement.Au cours d’une année, la période de végétation (phénophase feuillée) n’est que de 4 à 6 mois et la productivité primaire se partage ainsi en :– une fraction qui meurt et tombe, surtout à l’automne, sous forme de chute de litière utilisée par des détritivores;– un apport aux consommateurs herbivores ou phytophages;– une exportation de matériaux divers, surtout importante lorsqu’il y a exploitation par l’homme (éclaircies ou coupes à blanc);– une contribution à l’augmentation de la phytomasse ligneuse: c’est l’incrément de la forêt souvent exprimé en mètres cubes de bois fort (jusqu’à 7 cm de diamètre) par les forestiers occidentaux. Cet incrément est de 6-13 t MS/ha, soit 3 à 8 m3 à l’hectare de bois fort pour le hêtre et environ 5 m3 pour le chêne, dont la densité est plus forte; pour des bois légers à croissance rapide comme celui de certains résineux, l’incrément peut être bien plus important: il atteint jusqu’à 20 m3 pour l’épicéa.La fraction de la production nette PN entrant dans le réseau de consommation par l’intermédiaire des herbivores et phytophages est généralement très faible; les vertébrés sont peu importants: 5-7 kg MS/ha pour les mammifères, 1-2 kg MS/ha pour les oiseaux. Les invertébrés peuvent être bien plus abondants, surtout lors d’années d’invasion par des chenilles défoliatrices qui affectent profondément la fixation d’énergie: leur biomasse peut alors passer de 10-20 kg MS/ha (valeur normale) à 75-150 kg MS/ha, ce qui correspond à une destruction de plus de la moitié de la production foliaire. L’entomofaune circulante, qui atteint une biomasse de 15-20 kg MS/ha, est composée d’insectes (surtout diptères) commençant leur vie à l’état de larve dans la litière et dans l’humus (rôles sapro-, rhizo- ou zoophages). Tous ces animaux n’utilisent qu’une partie de la phytomasse consommée à édifier leur biomasse (productivité secondaire); le reste est perdu sous forme de fèces et d’énergie respiratoire.La chaîne des décomposeurs l’emporte totalement sur celle des consommateurs. En effet 50 à 75 p. 100 de la productivité primaire nette donnent chaque année une litière qui entre dans le réseau de décomposition. De cette litière une fraction, lente à se décomposer, subsistera pendant un nombre plus ou moins grand d’années, s’ajoutant à la litière préexistante; une autre fraction, livrée aux détritivores et aux transformateurs, deviendra de l’humus; le restant sera reminéralisé (recyclage) par des bactéries minéralisatrices.La biomasse des bactéries n’est que de quelques kilogrammes de matière sèche par hectare, mais leur productivité est particulièrement forte, car elles ne cessent de se multiplier en même temps qu’elles manifestent les activités les plus diverses – décomposition, nitrification, dénitrification, fixation de l’azote, reminéralisation. Les animaux décomposeurs de la pédofaune sont avant tout des bactério et mycophages pâturant les peuplements de micro-organismes développés dans et sur la litière.Forêts de résineuxLes grandes forêts boréales toujours vertes de l’hémisphère Nord, adaptées à des climats souvent difficiles, n’ont en zone de taïga qu’une productivité médiocre. Rodin et Basilevic donnent, pour les forêts de l’U.R.S.S., les valeurs moyennes suivantes :– biomasse en tonnes par hectare = 100 en taïga du Nord, 260 en taïga centrale et 330 en taïga du Sud;– productivité nette par hectare et par an, respectivement 4,5 t, 7 t et 8,5 t.Cependant, la productivité varie fortement avec l’âge et, dans la taïga du Sud, elle peut passer par un pic de 20 t/ha/an pour les forêts en pleine croissance sur bon sol, la moyenne étant de 14 t/ha/an.L’épicéa a été et est encore aujourd’hui la principale essence de reboisement des zones peu fertiles de l’Europe occidentale. Ici, protégées par l’homme, les plantations monospécifiques d’épicéas (écosystèmes culturaux) réagissent bien mieux à un climat plus favorable que celui de leur pays d’origine, et leur production nette atteint 15-20 t/ha/an, même sur des sols médiocres.Les pins (Pinus) , grâce à leur grande frugalité, colonisent des sols très pauvres. Ainsi, la principale «usine à bois» de l’Europe occidentale est la forêt de pins maritimes (Pinus pinaster) rétablie dans le triangle des landes de Gascogne. Entre 24 et 28 ans, l’accroissement des troncs atteint de 5 à 20 m3/ha et la production globale est de l’ordre de 14 t/ha/an.Forêts tropicalesLes forêts tropicales, particulièrement les forêts tropicales humides (forêts sempervirentes ou «rain forest»), sont caractérisées par la luxuriance de leur végétation dans laquelle se superposent une série de strates; les arbres les plus hauts peuvent dépasser 50 m.Une solide étude d’écosystème forestier tropical humide a été réalisée en Malaisie à Pasow. La biomasse épigée dépasse 430 t (matière sèche) par hectare, dont 346 t pour les troncs et moins de 8 t pour les feuilles. La production primaire nette peut être estimée à 26,7 t/ha/an, dont 13,7 concernent les parties ligneuses et 7,4 les feuilles. La respiration de ces végétaux est intense, par suite de la température constamment élevée, même la nuit: elle représente l’équivalent de 50,5 t/ha/an, ce qui permet d’estimer la production brute de l’écosystème à 77,2 t/ha/an. L’écart considérable entre production brute et production nette est caractéristique des milieux tropicaux, et tout spécialement des forêts sempervirentes.Sur cette production, la quantité consommée vivante est excessivement faible: de l’ordre de 0,3 t/ha/an seulement, ce qui signifie que l’essentiel de la matière vivante formée est utilisé après sa mort par des détritivores et plus encore par des micro-organismes décomposeurs. Quant à la grande faune des forêts, si elle peut être très diversifiée, elle n’est représentée que par de très faibles densités d’individus de chaque espèce. De plus, elle est en grande partie dans les strates supérieures (singes, oiseaux), là précisément où se trouvent les feuilles vivantes qui sont le point de départ des chaînes trophiques.ToundrasTout à l’opposé des forêts tropicales, les toundras sont des formations végétales très basses localisées dans les régions les plus froides du globe. Les formes végétales dominantes y sont des lichens ou des mousses.La production annuelle est très faible, inférieure à 3 t/ha/an, et le temps de renouvellement de la biomasse relativement très long pour des plantes herbacées de petite taille. La vitesse de décomposition est également très lente.La vie animale, surtout endogée, et pour une part aquatique, dans un sol gelé une grande partie de l’année, mais imbibé d’eau, est représentée surtout par des détritivores dont l’action est complétée par celle des micro-organismes décomposeurs qui ont, par suite du froid, une action très ralentie. De fait, le cycle saisonnier est très accentué et marqué par une longue phase d’inactivité hivernale. Quelques vertébrés supérieurs peuvent survivre, à la faveur de l’homéothermie, mais leur importance quantitative est très faible, qu’ils soient herbivores ou carnivores.Prairies des zones tempéréesContrairement aux écosystèmes forestiers, la prairie est destinée à développer la productivité secondaire des grands herbivores. Relativement appréciable, celle-ci sera «exportée» sous forme de viande, de lait, de cuir, de laine. Ce sont pourtant encore ici les décomposeurs et reminéralisateurs (lombrics, bactéries, champignons) qui profitent le plus d’une productivité primaire où la part des racines est très importante.Cette prairie peut devenir permanente, ou faire partie d’un système de rotation dans lequel elle est périodiquement retournée pour faire place à des cultures. La récolte de l’herbe – surtout graminées et légumineuses – se fait de deux façons: la fauche (prés à foin) ou la dent des animaux (pâtures), ou parfois des deux combinées.La productivité primaire dépend du nombre de fauches ou du nombre de passages du bétail; en fonction de ce nombre, elle augmente d’abord, pour diminuer ensuite (surpâturage destructeur). Elle est en général de 5 à 15 t MS/ha/an pour les parties aériennes, mais irrigation et engrais peuvent, dans quelques cas, l’amener à 25 t. La productivité des racines est difficile à déterminer, celles-ci se renouvelant tous les 1, 2, 3 ans ou plus, selon les espèces; de plus, les racines sont des organes régulateurs, participant à la repousse de l’herbe après chaque coupe, grâce aux matières stockées pendant les intervalles. La phytomasse des racines est de l’ordre de 10 t MS/ha et l’on peut en estimer la productivité à 5 t/ha ou même davantage.Les animaux pâturants ne sont sur le terrain que pendant la période de végétation active (180-225 j/an); leur productivité dépend de la quantité et de la qualité de l’herbe (digestibilité, protéines digestibles, oligoéléments); elle dépend aussi du nombre de passages (division de la prairie en parcelles). On a établi que, dans de bonnes conditions, une vache laitière de 500 kg mangeant 70 kg d’herbe (14 kg MS par jour) produit 15 l de lait par jour; un bœuf à l’engrais de 350 kg accroît son poids vif de 1 kg/j en mangeant 45 kg d’herbe (9 kg MS) par jour.Le rendement et la saison de pâturage peuvent être augmentés par l’utilisation de foin et d’aliments plus concentrés. Ces aliments fournis à l’étable sont souvent produits à la ferme par des écosystèmes annexés à la prairie permanente: prés de fauches, luzernières, cultures de céréales ou plantes à tubercules, etc. Cela nous introduit dans un type d’écosystème plus complexe, l’agroécosystème.AgroécosystèmesAu sens large, l’agroécosystème est le plus ancien écosystème étudié et mis en place par les hommes du milieu rural. Une fois réalisée la domestication des plantes et des animaux, il devenait vital pour les paysans d’obtenir les meilleurs rendements de leur terre, en assurant les assolements nécessaires, la conservation de l’humus et le recyclage des nutriments minéraux par l’équilibre agro-pastoral .La forêt joue aussi un rôle dans la valorisation des terres agricoles (matériaux divers, bois de feu, haies pare-vent, pâturage sous forêt, etc.), de sorte que l’on peut parler, avec Kuhnholz-Lordat, d’équilibre sylvo-agro-pastoral . Il s’agit de maintenir, suivant les régions, des proportions équilibrées écologiquement de forêts (silva ), de pâturages (saltus ) et de cultures (ager ); la localisation topographique peut être aussi très importante: les forêts sur les crêtes régularisent le cycle de l’eau et empêchent l’inondation des vallons, où les apports réguliers d’alluvions fertiles forment des sols favorables aux cultures; les pentes, où le travail de la terre est plus difficile, peuvent servir de pâturages, à condition d’en éviter la surcharge.On voit apparaître ici les notions de paysage écologiquement fonctionnel, d’écopaysage, voire d’écorégion.Ayant envisagé déjà les cas de la forêt et de la prairie pâturée, nous considérerons ici l’agroécosystème dans son sens étroit de champ cultivé. Une partie très importante de la productivité primaire est exportée, non seulement par la récolte des grains, tubercules ou autres organes recherchés, mais souvent aussi sous forme de paille, feuilles ou déchets divers servant à l’alimentation animale, à la litière d’écurie, au compostage, etc. C’est dire que la restitution au sol de matière organique et de nutriments minéraux est faible, ce qui nécessite une importation compensant les matières premières perdues si l’on veut éviter une dégradation et un appauvrissement progressif du sol; aujourd’hui, le traditionnel fumier fait une place de plus en plus grande aux engrais chimiques.Pour fixer les idées, la productivité primaire nette d’un bon champ de blé, sur sol sablo-limoneux fertile, est de 16,7 t MS/ha, dont 6,5 t de grains, 2,2 t de glumes et glumelles, 5 t de tiges, 2 t de feuilles, 1 t de racines. Les 6,5 t de grains et 7,5 t de «pailles» (glumes, tiges et feuilles séchées) sont exportées; 2,7 t de bas de tiges, racines et feuilles sont restituées, sous forme d’éteule abandonnée sur le terrain après la coupe.Dans les pays développés, la productivité des cultures est très fortement augmentée par l’importation de grandes quantités d’énergie fossile (pétrole, gaz naturel) nécessaire (machines agricoles, transports, irrigation, engrais, pesticides, aménagement des installations, etc.).L’exemple de l’écosystème «champ de maïs» aux Etats-Unis, étudié par Pimentel et ses collaborateurs (1973) montre que, en moyenne, le champ de maïs reçoit, pendant la période de végétation, 5,1.109 kilocalories d’énergie solaire totale, par hectare; 1,26 p. 100 de cette énergie est convertie en production (PN1), de maïs total, soit 64 360.103 kcal, correspondant à 16 t MS/ha/an; environ le tiers de cette production correspond aux grains: 20 432.103 kcal (5,1 t MS grains/ha/an), représentant en tout environ 0,4 p. 100 de l’énergie solaire reçue (fig. 6). Une telle productivité ne peut être obtenue qu’avec l’intervention, à de très nombreux niveaux, de quantités importantes de combustibles fossiles:– énergie de fabrication des engrais (h), surtout azotés: en tout 2 641.103 kcal/ha;– énergie de fabrication des pesticides (j);– énergie d’irrigation (i): en moyenne, 85.103 kcal d’énergie sont nécessaires pour amener l’eau et la distribuer sur 1 hectare de culture de maïs;– énergie de séchage car le grain mûr de maïs hybride qui contient 13 p. 100 d’eau risque de moisir et il faut utiliser 300.103 kcal d’énergie pour le sécher;– énergie électrique pour les besoins les plus variés de la ferme (b, c): 775.103 kcal pour 1 hectare de champ cultivé;– apport de graines sélectionnées (maïs hybride) pour plantation, soit une importation d’énergie dans le système de 157.503 kcal;– énergie humaine (d): aux États-Unis où un ouvrier agricole travaille 40 heures par semaine en consommant 21 770 kcal/semaine et où il passe, pour toute la saison, 22 h sur 1 hectare de champ de maïs, cela représente 12,3.103 kcal d’énergie alimentaire.Ces divers apports d’énergie représentent plus du tiers de l’énergie solaire fixée sous forme de grains (1 kcal d’énergie fossile pour produire 2,8 kcal de grains). Dans les cas, très fréquents, où les grains de maïs servent à l’alimentation des animaux domestiques, il faut dès lors compter 3 kcal d’énergie fossile pour produire 1 kcal de viande par l’intermédiaire de 9 kcal de grains.Si l’on considère, par ailleurs, que l’alimentation humaine est à la fois d’origine végétale et d’origine animale, c’est-à-dire que l’homme se situe à la fois au niveau des herbivores et à celui des carnivores et qu’entre ces deux niveaux la perte d’énergie est de 100 à 1, on voit d’après l’exemple précédent quelle économie énergétique peut représenter pour l’humanité de demain un régime de plus en plus végétarien. De fait, les populations qui doivent vivre à des densités considérables, comme c’est le cas des habitants des deltas indochinois par exemple, ont depuis toujours une alimentation essentiellement végétale.3. Contrôle du fonctionnementÀ l’inverse d’un organisme, un écosystème ne possède pas d’organes de contrôle dont le rôle serait analogue à celui des systèmes neuro-endocrinien et immunitaire des vertébrés, dont la fonction est d’assurer à la fois l’intégration du fonctionnement des organes effecteurs, la constance du milieu intérieur et l’adaptation optimale aux conditions changeantes de l’environnement. L’écosystème n’est donc pas une sorte de «superorganisme» installé dans un ensemble de conditions physiques qui lui conviennent. La cohésion de sa composante vivante est entièrement assurée par la complémentarité d’intérêts (en termes de valeur adaptative, ou fitness ) des espèces qui la composent. Celles-ci sont associées sous l’effet de deux types de contraintes, les unes externes, les autres internes.Les contraintes externes constituent ce que l’on appelle couramment les facteurs limitants du système, dont ils déterminent à la fois les limites spatiales et la structure temporelle; ils influencent aussi le taux de renouvellement de ses composantes vivantes. L’identification des principaux facteurs limitants des écosystèmes est ancienne, bien antérieure à la naissance de l’écologie en tant que discipline autonome. Dans les écosystèmes terrestres, les rôles capitaux de la température et des précipitations, de l’altitude et de la latitude, ont été reconnus dès le début du XIXe siècle; les lignes isothermes, les isohyètes et les courbes de niveau interviennent à ce titre dans tous les travaux des biogéographes. La lumière ou la teneur en oxygène et en sels minéraux jouent un rôle essentiel dans les milieux aquatiques. La restriction spatiale et l’isolement géographique ont depuis longtemps été impliqués en tant que variables influençant la richesse spécifique et par conséquent la structure trophique des biocénoses.L’analyse des modalités d’action de ces facteurs limitants exogènes est cependant bien loin d’être achevée. On a, par exemple, trop utilisé jusqu’ici les moyennes annuelles de la température ou des précipitations, et trop souvent sous-estimé les valeurs extrêmes. Quelle que soit la latitude, en effet, les variations saisonnières des paramètres climatiques majeurs ont de profondes répercussions sur la structure et la dynamique des écosystèmes. L’importance des variations journalières des mêmes variables n’a pas reçu non plus toute l’attention qu’elle mérite, en particulier sur les hautes montagnes tropicales. Dans les phénomènes d’insularité, l’importance de la distance des sources possibles de migration et des phénomènes d’extinction spontanée n’a été reconnue que très récemment.À côté de ces contraintes exogènes, la part des contraintes endogènes est difficile à apprécier dans un écosystème naturel «mature» dont l’homéostasie est généralement excellente. En revanche, dès que l’on considère des successions dans le temps (séries biocénotiques progressives ou régressives) ou certaines situations expérimentales (agrosystèmes), il devient possible d’entrevoir l’importance de certains mécanismes internes de régulation. Les uns ont pour fonction de limiter la taille des populations spécifiques, de façon à éviter qu’elles ne dépassent la capacité de charge du milieu: tel est le cas des facteurs socioécologiques de régulation de la natalité et de la mortalité dépendant de la densité, dont l’existence semble certaine chez un certain nombre de vertébrés. Quand ces facteurs ne jouent plus, généralement du fait d’une intervention humaine, une pullulation de consommateurs peut aboutir à la destruction de leur propre habitat: la déforestation quasi totale de certains parcs nationaux africains consécutive à une surpopulation par les éléphants est un exemple connu de tous.D’autres mécanismes internes de régulation sont interspécifiques et ont pour effet d’empêcher ou de limiter la compétition à un niveau trophique donné. Telle paraît être la fonction de ces substances chimiques sécrétées par beaucoup de végétaux et qui peuvent empêcher la croissance d’espèces concurrentes (phénomènes d’antibiose et d’allélopathie). Certains agents pathogènes animaux (des virus aux hématozoaires) qui ne causent que des accidents mineurs chez leurs hôtes habituels, mais peuvent entraîner une mortalité très élevée chez des formes voisines non sympatriques, ont peut-être un rôle écologique voisin (myxomatose, herpès simien, certains trypanosomes, etc.). Des phéromones animales, en particulier celles exerçant un pouvoir répulsif, sont peut-être à ranger aussi dans cette catégorie.
Encyclopédie Universelle. 2012.